 Как обшить баню внутри вагонкой своими руками — пошаговая инструкция и советы экспертов Rodno
Как обшить баню внутри вагонкой своими руками — пошаговая инструкция и советы экспертов Rodno  Памяти папы
Памяти папы  "Фейри файр": ярмарка чудес и замечательные животные))
"Фейри файр": ярмарка чудес и замечательные животные))  Хайфа
Хайфа  Зелебобик наш?
Зелебобик наш?  Китай. Опять.
Китай. Опять. 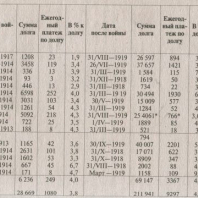 Долги воюющих стран
Долги воюющих стран  Лиссабон - Пантеон и нижняя Альфама
Лиссабон - Пантеон и нижняя Альфама  Ещё один подавился деньгами
Ещё один подавился деньгами
14.11.2022 —
Мысленное путешествие во времени
Мозг, как сигнальная система.
 evan_gcrm — 26.11.2022
evan_gcrm — 26.11.2022

Попытка продемонстрировать, что принцип становления молекулы в самоорганизуемую сигнальную систему на основе физических свойств, лежит в основе формирования как нервной системы, так мозга.
Мозг, как объединение различных систем изначально формировался как рецепторная сигнальная система. По своей сути это молекулярный 3D(трехмерной пространственной упаковкой систематизированной/оформленной информацией) белковый датчик, фиксирующий изменение определенного параметра.
Возможно, первым параметром была температура и скорость сборки протомолекулы. Трехмерность длинных белковых цепей естественным образом формирует такие пространственные положения, что в определенных частях упакованной цепи могут возникать структуры, похожие на каналы. Именно в них возникают ионные токи. Случайное перераспаковывание цепи приводит к сбросу концентраций локально накопленных ионов. Таким образом более стабильная молекула получает приоритет по длительности существования.
Возможно, способность создавать ионные токи на определенных участках белковой молекулы-рецептора спровоцировала необходимость создания интерпретаторов этих новых сигналов. И как в следствии его фиксации и кодирования. Результатом явилась способность к коммуникации с другими молекулами путём обмена концентрациями а потом и потенциалами.
Таким образом, любая молекула становится самоорганизуемой сигнальной системой - способ упаковки и расположение в пространстве становятся атрибутами «успешности» выживания/времени существования молекулы. Более сложно пространственно организованная/упакованная цепь формирует новые феномены (ещё и электрические, вдобавок к химическим). В свою очередь феномены и аппарат измерения изменяющихся состояний создаёт потребность в кодификации/структурировании качественно нового потока данных. В этот момент молекула/мембрана приобретает функции интерпретатора команд. Ионный ток формирует первичное поле молекулы/мембраны, являясь источником ресурса и качества его расходования.
Протонные помпы именно об этом.
Наличие электросенсорных качеств позволило увеличить количество атрибутов окружающих молекулу систем. То есть преимущество приобретают те изомеры/клоны молекулы-рецептора, которые имеют более длительное время существования или легко встраиваются в более сложные информационные системы (мембраны, как кооператив протонных помп), получая таким образом пролонгацию своего существования за счёт усложнения системы (много каналов) и усложнения формата данных.
Конкурентное преимущество получают молекулы/кооперативы протонных помп которые начинают кодировать событие/возмущение поля/флуктуацию пространственно временного континуума разноформатными данными. Вслед за концентрацией ионов (кислотность среды) и молекулы/мембраны начинают дифференцировать изменения поля.
Возможно, именно это свойство возникло как ответ на естественную причину - молекулы могут формировать поле и реагируют на поля вокруг. Естественно, мембраны, получившие возможность использовать свои дипольные качества, сформировали систему обработки и фильтрационного реагирования на электромагнитные возмущения в широком смысле. Таким образом последующая дипольность нейрона получила уже сформированный корпус правил поведения рецепторов электроактивности. Трудно определить степень «оживленности»/наличия жизни у таких молекул/мембран - самосборка, вероятно, была возможна без механизма днк/рнк и дуплицирования/наследования.
Физические свойства скорее, чем химические, определяли первичные предпочтения молекул.
Их энергетический статус, как способность вступать в новые комбинаторные соединения или выступать в качестве катализатора/провокатора несвязанных, сторонних объектов приносят выгоду более «коммуникативным» молекулам. То есть для каждой молекулы становилось необходимым интерпретировать не просто изменение градиента концентрации в бинарном понимании (+/- ; да/нет). Мембраны получили третье состояние (ноль) информации, которое описывает середину между максимумом заряда на мембране и минимумом реполяризация. Это радикально иные характеристики системы. При наличии двух сигнальных систем такой рецептор получает фазовый переход - способность управлять энергией и манипулировать своим состоянием через временную фиксацию. Эволюционно гораздо позже, способность манипулировать объектами снаружи станет ключевым фактором в конкуренции молекул и выживания наиболее адаптированных.
Просто приспосабливаться становится недостаточным.
Протомолекула выбирает случайным образом из всего поля возможностей (температура, заряд, кислотность) данные и выдаёт ограниченное пространство решений по самоупаковке в пространстве. Мозг как орган и система формировался как пространственно/морфологически так и функционально для реализация потребности анализа метаморфных данных, которых с добавлением электро-магнитной составляющей стало больше. Кодирование и обмен усложнились, но позволяло объединяться с похожими по функционалу, при условии что новое агрегированное состояние отвечало двум требованиям: энергоэффективности и ресурснопозитивной (даже краткосрочно) зависимости пространственной структуры протомолекулы от формата волнового взаимодействия.
Наличие аппарата управления информационным/сигнальным потоком в сочетании со способностью агрегировать/управлять ресурсными ионными/протонными потоками явилось предусловием в необходимости обмена информацией.
То есть первичные рецепторы могли вполне интенсивно эволюционировать на домембранном этапе, находясь в состоянии отдельных субьединиц и склеиватся в конгломераты.
Начиная с примитивных молекул-прогениторов (глюкоза, глутамат) ранние нейроны уже имели весь функционал мембранных рецепторов.
Уже на этом уровне развития мозга/нервной системы мембраны нейронов умеют не только перераспределять течения электронов, они умеют управлять другими клетками, вызывая сокращения и движение.
Чтобы стать сенсором и начать получать кванты информации извне, молекула должна быть резонансной получаемому сигналу. Молекула-резонатор является действующим прототипом всех сенсорных систем. Такая молекула принимает сигналы но сама может оставаться условно «бесшумной».
Первичное кодирование - перевод тепловой энергии в химическую связь (или волновое воздействие в конформационное переформатирование). Таким образом, в ходе наблюдения за последовательностью трансформации молекул наблюдается трансформация сигнальной системы и языка в частности, как логического описателя всех событий внутри молекулы.
Под языками мозга понимается набор символов-сигналов которыми оперируют колонии высокодифференцированных нервных клеток в сочетании с описанием операций и структуры всех типов манипуляций внутри системы. В процессе развития нервная система интегрировала и ассимилировала множество химических и электромагнитных языков обмена информацией (химические, электро-магнитные и волновые).
Именно языки обработки информации вынуждали систему меняться морфологически, т.е. добавление нового морфологического признака следовало за потребностью системы в расширении.
Первичные языки полимерных цепей, дошедшие до нас в виде внеклеточных форм жизни (вирусоиды, полипептиды, вирусы), позволяют оценить из какого субстрата развивалось кодирование «смыслов». Вариантов было несколько и то, что до нас дошла единая форма позволяет судить о ее конкурентном преимуществе, универсализме и пластичности языка передачи информации.
Преимущество заключалось в лаконичности и компактности кода.
Фактически, четыре символа, обьединенных по фазе/вектору/резонатору(сенсорной системы) позволяют описать все события внутри системы и в окружающей среде (фиксированные системой). Причем, код настолько универсальный, что позволяет добавлять новые функции путем «перепрошивки» старой конструкции молекулы (содержащей реестр всех предыдущих изменений самой молекулы и событий/действий в окружающей среде).
Возможно, подобное накопление информации явилось прообразом памяти.
Все базисные языки описывают самые древние молекулярные взаимодействия и по происхождению восходят к молекулам-резонаторам (сенсорам). Изначально, молекуле необходимо идентифицировать/определить сигнал в сигнальном потоке. Обычно, в простейших бимолекулярных системах таким сигналом является предпорговое/надпороговое изменение стандартных факторов, влияющих на молекулу (фотоны/нейтроны способные выбивать электроны с внешних орбит крайних участков полимерных цепей или изменение градиента кислотности/щелочности вызывающее скручивание полимерной цепи. Термический фактор или волновое воздействие).
Вероятно, одними из первых морфологических элементов должны были сформироваться резонаторы, как прообразы будущих сенсоров.
Поэтому, сигнальная функция нервной системы является первостепенной.
Для возникновения первичной сигнальной системы требуется:
• пространственное положение атомов скрученной цепи по отношению к другим участкам.
Одно из преимуществ кооперации полимерных молекул - множественные пространственные положения и множественные места контакта разнородных участков внутри клубка в состоянии сворачивания, позволили совмещать новые сенсоры/рецепторы без конфликта для всей молекулы, но с сохранением базовых свойств всех компонентов системы. Внутри одного конгломерата способность различать отличия позволяла создать систему свой/чужой и взаимосвязано развивать сенсорные/сигнальные элементы системы, совмещая разные виды резонаторов. Так формировалась селективность.
Сложность трехмерных цепей подводит такую скрученную цепь к сложному выбору стратегии: либо продолжать накапливать ресурс в виде присоединения все новых блоков со своим набором сигнальных участков, либо получить преимущества не от накопления массы энергии/информации, а от качества принятой информации и систематизации способов использования приобретаемых функций.
Действительно, для удлинения цепи требуется небольшой набор операций. Быть длинными со всем допустимым множеством сенсорных/детекторных/резонаторных участков/фокусов можно было лишь ограничившись присоединением нового участка. Вторая стратегия предполагает энергоэффективность всей молекулы, которая достигается, путем уменьшения повторяющихся одинаковых участков и возрастанием избирательности сигнальных участков случайного набора сенсоров. В результате, более селективные молекулы стали сильнее во всех смыслах, чем их длинномерные конкуренты.
Усложнение первичных молекул возникло как следствие увеличения количества новых блоков в системах, способных масштабироваться. Такая система стремиться к расширению пространства вокруг в поисках новых элементов информации, которая поступает по мере освоения ресурсной или энергетической базы. Усложненная система, вероятно, имела более пролонгированные стабильные/устойчивые состояния в сравнении с угнетенными системами, которые не справлялись с состоянием возбудимости (переизбыток одного из внешних факторов разрушал молекулу). Следствием этого является деградация - сначала «идеологическая» (уменьшается количество возможностей взаимодействия с информацией и степеней свободы) и затем следует физическая деградация - распад молекулы на безсвязные логически атомарные единицы, которые с удовольствием используются для строительства в более успешных системах.
Со временем такая система переходит в состояние динамического равновесия. Но постоянное изменение условий окружающей среды (жесткое гамма-излучение ранних этапов сменялось на жесткое ультрафиолетовое, радикально изменялись температурные режимы, с появлением дипольного заряженных молекул изменялась природа электрических взаимодействий, кислородная катастрофа) не дает системе возможности стабилизироваться. Постоянно возникает дефицит/перенасыщение, в системе, стремящейся к равновесию и в то же самое время, вынужденно находящейся в перманентом динамическом поиске равновесия.
Первичные молекулы формировались в «домембранные» времена и возможно, именно первичная электрохимическая кооперация молекул и послужила первым стартовым процессом начала сборки мембран.
Форма днк/рнк была предопределена химией и физикой взаимодействия первичных полимеров. В тот же момент появилась еще одна характеристика системы (новая информационная категория, которая должна была описывать для системы пространственное положение каждого компонента). Существование молекул в более сложном, чем двумерная изомерия, формате позволяло улучшить «выживаемость» скрученной полимерной цепи - при критическом/фазовом переходе такая система имела более высокие шансы на продолжение существования, за счет повторения свойств всей цепи в неповрежденых участках. Повреждение части цепи не приводило к разрушению всей цепи, что стало преимуществом перед молекулярными конструкциями более простой организации и без пространственных изменений. Гораздо позже скрученная цепь научится саморегулировать свои состояния - самособираться (как ответ на изменения) и самораспускаться в двумерную форму в периоды условной стабильности/благополучия.
Пространственное положение атомов в скрученной цепи становится новой модальностью.
По мере увеличения удельной атомной массы полимеризированых цепей усложнялся и аппарат приема сигнала. За счет унифицированности всей цепи полимера достаточно одного изменения - присоединение еще одного атома/создание еще одной электронной пары с любого конца и вся цепь начинает владеть новым свойством. Такое присоединение и обладание новым качеством позволяет получать еще больше информации от окружающих систем.
Усложнение пространственной геометрии конгломератов скрученных/развернутых цепей создало набор вероятностных возможностей для этих клубков. Обьединение цепей в клубки происходило по принципу объединения в центральной части субъединиц с высокой долей общности/гомогенности компонентов, вынося пространственно на поверхность «инородные» концевые молекулярные находки всех участников кооператива. Таким образом формировалось множество клубков с одинаковыми (или сходными по строению цепи) внутренними блоками и «щетиной» концевиков на поверхности. Подобным образом сложные полимерные клубки научились определять сложность соседей и демонстрировать свои особенности - концевые выпячивания на собственной поверхности клубка. При наличии низкоуровневой молекулярной системы свой-чужой клубок получил функцию - поведение, как совокупность донорства информации и ее акцептирования. Уже на этом этапе незначительное (по меркам даже клетки) формирование в виде нестабильных полимерных сгустков получает очередной навык - обороны от «нежелательных гостей» - соединений с токсичными концевиками, путем отсутствия химического/электронного контакта или путем создания барьера из своих концевых окончаний, которые могут быть токсичны для соседей. Второй навык, нераздельно связанный с первым - способность присоединять к себе соседние полимерные образования, в т.ч. без их «согласия», просто за счет большей массы и длины цепи. Эти особенности включают разные сенсорные умения, которые реализовывались благодаря различным вариантам сборки полимерной цепи на предыдущих этапах.
Поведение, как совокупность ответов на поступающую информацию, появилось как адаптационная реакция сложнорганизованных молекул на наличие обилия сенсорных элементов внутри скрученной цепи. Поведение позволило молекулам манипулировать другими посредством обмана или делясь уникальными артефактами из собственной копилки сенсоров и сигналов.
Частотные характеристики молекул характеризуют скорость переключения из одного состояния в фазово иное. Однако, до-мембранное время это и неритмичное время или время множества разрозненных ритмов.
Селекция по ритмическим параметрам - зарождение Синхронизации.
Стоит упомянуть, что любая корпускулярная система находится под воздействием волновых эффектов и под влиянием полей, которые существуют стабильно и возникают спонтанно в результате взаимодействия всего множества частиц внутри полимерной цепи.
Языки мозга имеют целеположения - оптимизировать (энергетически, логистически, функционально) деятельность системы (не клеток), сформировать наиболее адаптивную и в то же время наиболее экспансивную (доминантную) адаптационную модель.
Стоить отметить, что язык мозга это не просто описание команд - где, как и что присоединить/удалить, это и распределенная база управления единицами информации.
Первичными информационным потоками для полимерных протомолекул являлись ионные потоки и физические каналы волновой и корпускулярной природы.
Язык упорядочивает систему, в которой он существует. Переводит внешние сигналы (заряд, пространственное положение, гравитация, ионизирующее излучение) в упорядоченную систему трансформации информации посредством изменения кода химической цепи.
По мере усложнения языка внутри системы усложняется и морфология носителя/интерпретатора.
Например, зачем нейроны имеют множество трансмиттеров, хотя, по факту всю нервную сеть можно сделать на одном виде сигналов, с одним трансмиттером?
Такое большое количество трансмиттеров было унаследовано в процессе коммуникации!
Нейротрансмиттеры были унаследованы и именно они создали нервную систему.
На результатах многочисленных экспериментов можно сделать выводы, что нервная система и нейроны появлялись в эволюции несколько раз и независимо. Это подтверждает тезис, что трансмитерная специфичность может быть связана с независимым появлением нейронов в эволюции.
У простейшего многоклеточного Placozoa нет нейронов, но они способны к разнообразному поведению - убеганию, поисковому и пищевому поведениям. Уже на таком уровне они имеют десятки трансмитерных систем, разные клетки имеют разные наборы трансмиттеров - именно на этом строится координация поведения этих клеток. Клетки знают как они должны отвечать на определенный сигнал в межклеточном пространстве. Это наследуемая функция и отбор с закреплением определенного спектра рецепторов для этого типа клеток. Но есть и пластичность, когда подстройка происходит во время развития. Нейротрансмиттеры создали нервную систему, объединяя предковые популяции секреторных клеток для поведенческой координации БЕЗ синапсов!
Нервная система унаследовала не просто трансмиттеры, она унаследовала систему клеточной интеграции - язык управления. Даже у относительно небольшого и медлительного организма, клетки уже умели договариваться, предсказывать будущее и быть адаптивными. Все это происходит на базе химической сигнализации - клетки были разной химической природы и имели разные спектры чувствительности к химическим веществам.
Что же добавилось с появлением в нейроне электричества?
Прежде всего скорость и увеличение протяженности передачи сигнала. Но система коммуникации и сам принцип управления нейрональным ансамблем остался прежним.

Сохранено
|
|
</> |
Оставить комментарий
Популярные посты:
Предыдущие записи блогера :
31.10.2022 —
Музыкальная пауза
Архив записей в блогах:
хочу задать читателям несколько вопросов и надеюсь они примут активное участие в ...
Сбываются самые мои худшие прогнозы. Криминально-олигархический режим в России, чтобы прикрыть сдачу Донбасса и Сирии, решил втянуть Россию в новую военную авантюру. На этот раз в Ливии. Именно такие мысли приходят в голову от загадочного визита т.н. командующего Ливийской ...
Декабрь, идёт дождь, летит время. Вот-вот запахнет мандаринами. Вчера серьёзный дяденька тащил ёлку. Тощую, с виду болезненную. Наверно, подобрал её, как подбирают бездомного котёнка. Принесёт домой, отмоет, высушит феном, вызовет хвойного ...
Культовый кинорежиссёр (1928-2022)... Родился в Амурской области, жил в Питере... Орденоносец, Народный артист РСФСР, обладатель премии "Ника"... Перечень шедевров зашкаливает. Среди них... 1966 — Начальник Чукотки 1969 — Мама вышла замуж 1970 — Семь невест ефрейтора Збруева 1972 — ...
Хорошая новость! Россельхознадзор своим письмом разрешил ввозить украинскую продукцию в Крым до конца года. За исключением свиней. Впрочем, как Путин говорил, Россия сама себя обеспечивает свининой. Да и кому нужна эта свинина? Я, например, ее не ем, наша фирма тоже свининой не торгует, от ...



